Строение днк бактерий. Днк бактериальная. Роль бактерий в природе
Носителем генетической информации бактериальных клеток является ДНК. Она представляет собой двойную спираль, состоящую из двух полинуклеотидных цепочек. ДНК сравнивают с винтовой лестницей и с двойным электрическим кабелем. Остов ДНК состоит из фосфатных групп и дезоксирибозы. Полипептидные цепи соединены между собой водородными связями, которые удерживают друг с другом комплементарные азотистые основания. Строение ДНК бактерий аналогично таковому клеток эукариотического типа (растений, животных, грибов). В отличие от бактерий у вирусов геном представлен одной нуклеиновой кислотой – ДНК или РНК. Бактериальные клетки, кроме ДНК, могут иметь генетически полноценные образования функционирующие автономно. Необходимо подчеркнуть, что носителями наследственности бактерий кроме ДНК являются плазмиды и эписомы. В этой связи, любая структура бактериальной клетки, способна к саморепликации, называется репликон, т. е. репликонами бактерий являются нуклеотид, плазмиды, эписомы. Плазмиды не связаны с нуклеотидом, они пребывают в цитоплазме клетки автономно, эписомы могут находиться в свободном состоянии, но чаще всего они реплицируются вместе с ДНК.
Бактериальная хромосома представлена одной двунитевой молекулой ДНК кольцевидной формы и называется нуклеотидом. Длина нуклеотида в растянутом виде составляет примерно 1 мм. Нуклеотид – эквивалент ядра. Расположен он в центре бактерии. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Нуклеотид можно выявить в световом микроскопе. Для этого надо окрасить клетку специальными методами: по Фельгену или по Романовскому-Гимзе. Электронно-микроскопическое исследование показало, что один конец ДНК прикреплен к клеточной мембране. Видимо, это необходимо для процесса репликации ДНК.
Выращивани бактерий в пробирке. Фото: Tess Watson
В отличие от клеток эукариот у прокариот отсутствуют митохондрии, аппарат Гольджи и эндоплазмотическая сеть.
Каждая нить ДНК состоит из звеньев – нуклеотидов. В состав нуклеотида входит одно из азотистых оснований (аденин, гуанин, тимин или цитозин) дезоксирибоза и фосфорная кислота. Приблизительно 1500 нуклеотидов составляют ген средней величины. Таким образом, ген представляет собой определенный участок ДНК, ответственный за проявление и развитие конкретного признака. Гены в ДНК расположены линейно, они дискретны, способны к саморепликации. Последовательность аминокислот в синтезируемом белке, определяется последовательностью нуклеотидов в гене.
С точки зрения функциональной гены подразделяют на структурные, регуляторы, промоторы и гены-операторы.
Структурные гены, представляют собой гены, обуславливающие синтез ферментов, участвующих в биологических реакциях и в формировании клеточных структур.
Гены-регуляторы ответственны за синтез белков, регулирующих обмен веществ. Эти гены могут влиять на деятельность структурных генов.
Гены-промоторы детерминируют начало транскрипции. Они представляют собой участок ДНК, который распознает ДНК-зависимый РНК-полимеразой.
Гены-операторы являются посредниками между структурными генами, промоторной областью и генами-регуляторами.
Совокупность генов-регуляторов, промоторов, операторов и структурных генов называют опероном. Следовательно оперон является функциональной генетической единицей, несущей ответственность за проявление определенного признака микроорганизмов.
Различают индуцибельные и репрессибельные опероны. Например, индуцибельным опероном является Lac-оперон, гены которого контролируют синтез ферментов, утилизирующих лактозу в микробной клетке. Если клетка не нуждается в лактозе, оперон поддерживается в неактивном состоянии и, наоборот.
Примером репрессибельного оперона может служить триптофановый оперон, обеспечивающий продукцию триптофана. Этот оперон обычно постоянно функционирует, а его белок-репрессор находится в пассивном состоянии. В случае повышения содержания триптофана в клетке аминокислота вступает в связь с репрессором и активизирует его. Репрессор ингибирует работающий оперон и прерывает синтез триптофана.
Важнейшее свойство ДНК – способность к репликации. Репликация может протекать по тета-типу и сигма-типу. Репликация ДНК по тета-типу начинается в определенной точке в виде «вздутия» и распространяется вдоль молекулы в двух направлениях, проходя через промежуточную структуру, напоминающую греческую букву тета. При этом типе репликации сохраняется одна из цепей исходной молекулы ДНК, а вторая синтезируется из нуклеотидов.
Репликация ДНК по сигма-типу осуществляется через промежуточную структуру, напоминающую греческую букву сигма, откуда и название этого типа. Этот тип репликации наблюдается в процессе коньюгации бактерий и некоторых фагов. При этом типе репликации происходит достраивание обоих нитей ДНК до двухцепочной ДНК.
Геном бактерий выполняет следующие функции:
· обеспечивает передачу биологических свойств по наследству;
· программирует синтез бактериального белка с определенными свойствами;
· участвует в процессах изменчивости бактерий;
· обеспечивает сохранение индивидуальности вида;
· детерминирует множественную устойчивость к ряду лекарственных веществ.
ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина,а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.[ ...]
Бактериальные ДНК - это высокополимерные соединения, состоящие из большого числа нуклеотидов - полинуклеотиды с молекулярным весом около 4 млн. Молекула ДНК представляет собой цепь нуклеотидов, где расположение их имеет определенную последовательность. В последовательности расположения азотистых оснований закодирована генетическая информация каждого вида. Нарушение этой последовательности возможно при естественных мутациях или же под влиянием мутагенных факторов. При этом микроорганизм приобретает или утрачивает какое-либо свойство. У него наследственно изменяются признаки, т. е. появляется новая форма микроорганизма. У всех микроорганизмов - прокариотов и эукариотов - носителями генетической информации являются нуклеиновые кислоты - ДНК и РНК. Лишь некоторые вирусы представляют собой исключение: у них ДНК отсутствует, а наследственная информация записана или отражена только в РНК.[ ...]
В бактериальных клетках в общем количестве оснований ДНК 32-65 мол.% гуанина и цитозина.[ ...]
Ядро бактериальной клетки. Примерно 1-2% веса сухой массы микроорганизмов приходится на ДНК, в которой заложена генетическая информация организма. У большинства микроорганизмов имеются области (или несколько областей), в которой сконцентрировано основное количество ДНК, имеющие определенную структуру (или органеллу) и называющиеся ядром. Ядро (или ядерное вещество) связано с цитоплазматической мембраной, независимо от того, окружено оно элементарными мембранами (как у амебы) или не имеет их (как у бактерий и сине-зеленых водорослей). Ядерное вещество активизируется в период размножения и при наступлении возрастных изменений, связанных со старением клетки.[ ...]
Сегмент ДНК (ген), который предназначен для молекулярного клонирования, должен обладать способностью к репликации при переносе его в бактериальную клетку, т. е. быть репликоном. Однако он такой способностью не обладает. Поэтому, чтобы обеспечить перенос и обнаружение клонируемых генов в клетках, их объединяют с так называемыми генетическими векторами. Последние должны обладать, как минимум, двумя свойствами. Во-первых, векторы должны быть способны к репликации в клетках, причем в нескольких копиях. Во-вторых, они должны обеспечивать возможность селекции клеток, содержащих вектор, т. е. обладать маркером, на который можно вести контрселекцию клеток, содержащих вектор вместе с клонируемым геном (рекомбинантные молекулы ДНК). Таким требованиям отвечают плазмиды и фаги. Плазмиды являются хорошими векторами по той причине, что они являются репликонами и могут содержать гены резистентности к,какому-либо антибиотику, что позволяет вести селекцию бактерий на устойчивость к этому антибиотику и, следовательно, легкое обнаружение рекомбинантных молекул ДНК.[ ...]
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками - гистонами - ив нуклео-иде расположена в виде пучка фибрилл.[ ...]
Применение методов рекомбинантной ДНК для получения биологических агентов для борьбы с загрязнениями находится на ранней стадии, но есть метод, который может оказаться полезным в обозримом будущем - это генетическое зондирование. Отбор организмов, способных трансформировать новое соединение, часто основан на способности использовать вещество как субстрат роста. Если рост слабый или субстрат только комета-болизируется, то методы селекции окажутся непригодными для идентификации деградативной способности. Следовательно, было бы полезно развивать генетическое зондирование для определения специфических последовательностей в плазмидах и хромосомах, это необходимо для определения катаболического потенциала, даже если этот потенциал не экспрессируется. Такие зонды разработаны для TOL-плазмид. Метод может определить одну бактериальную колонию, содержащую TOL-плазмиду, среди 106 колоний Escherichia coli. Такой мощный инструмент будет иметь огромное значение для выделения скрытых катаболических функций .[ ...]
Разработка изящной методики «клонирования» ДНК для получения большого количества точных копий специфических фрагментов ДНК (рис. 13.4) открыла в последнее время новые горизонты в изучении структуры, организации и функции генома. Если расщепить двухцепочечную ДИК одним из ферментов «рестрикции» (одной из нуклеаз), специфично узнающих и расщепляющих короткие последовательности нуклеотидов (4- 6 пар), то возникают в высшей степени воспроизводимые фрагменты ДНК. Концы двух цепей ДНК обычно бывают смещены относительно друг друга вследствие специфичности мест разрезания двухцепочечной молекулы, цепи которой комплементарны по составу оснований. ДНК обычно встраивают в плазмидный ген, важный для селекции, такой, как ген устойчивости к антибиотикам, что позволяет содержащим такую плазмиду бактериям расти в присутствии антибиотика.[ ...]
В бактериях при репликации образуется много копий плазмид, и таким образом можно «вырастить» большие количества встроенных фрагментов ДНК, а затем снова просто выделить их путем расщепления тем же самым ферментом рестрикции с разделением полученных продуктов гель-электрофорезом. Использование этого метода рекомбинации ДНК произвело революцию в изучении генов.[ ...]
Недавно было обнаружено, что мутагенное действие на бактериальные-ДНК-вирусы оказывают лучи с длиной волны 320-400 нм (область, близкая к зоне видимого света), обладающие низкой интенсивностью . Возможное влияние радиации в этом диапазоне длин волн на вирусы растений пока не обнаружено.[ ...]
Кривые зависимости реассоциацни от СОТ, полученные для бактериальной ДНК, лишены перегибов, а ДИК эукариот реас-социирует по другому типу (рис. 13.2). При низких концентрациях ДНК и коротком времени инкубации реиатурирует заметная доля одноцепочечной ДНК, а при увеличении СОТ образуется дополнительное количество двухцепочечных молекул, так что получается двухфазная кривая. Быстрая реиатурация при низких значениях СОТ показывает, что какие-то последовательности у эукариот повторяются много раз, т. е. до 10 000 раз и более.[ ...]
Отсутствие СХС может иммитироваться и в тех случаях, когда ДНК тестерных фагов не содержат сайтов, узнаваемых существующей в исследуемом штамме рестриктазой. Это явление представляет собой один из вариантов эволюционных адаптивных изменений бактериальных вирусов, призванных способствовать преодолению ими барьера СХС. Действие давления отбора в данном конкретном случае выражается в статистическом достоверном уменьшении числа или даже полной элиминации в фаговой ДНК последовательностей нуклеотидов, являющихся субстратом рестриктаз, характерных для клеток-хозяев бактериального вируса .[ ...]
Линдегрен описал возможные стадии образования бактериофага из ДНК профага, предположив, что профаг возникает как фрагмент чужеродной бактериальной ДНК, случайно проникший в клетку, который на ранних стадиях делится синхронно с бактериальной ДНК. Следующим важным этапом в развитии вируса явилось бы такое изменение профага, в результате которого стало возможным его независимое от ДНК клетки-хозяина размножение; в результате профаг использовал бы. все доступные нуклеотиды, нарушив тем-самым рост клетки-хозяина. Наконец, на какой-то более поздней стадии могла бы образоваться защитная белковая оболочка и возникли другие белки, что должно было обеспечить выживание ДНК вне организма хозяина и эффективное заражение новых клеток. Отделившийся фрагмент бактериальной ДНК вначале, очевидно, кодировал белки, приспособленные к бактериальным функциям. Необходимы очень существенные изменения в ДНК, чтобы могли возникнуть объекты настолько сложные и специализированные, как, скажем, фаг Т2 Е. coli, содержащие к тому же основания, которые в бактериальной ДИК отсутствуют.[ ...]
Генетическая информация бактерий не ограничивается ДНК, расположенной в нуклеоиде бактериальной клетки. Как уже отмечалось в предыдущих разделах книги, носителями наследственных свойств служат также внехромосомные элементы, получившие общее название плазмид. В отличие от ДНК ядерных эквивалентов-нуклеоидов, являющихся органоидами бактериальной клетки, плазмиды представляют собой независимые генетические элементы. Потеря плазмид или их приобретение не отражается на биологии клетки (приобретение плазмид оказывает положительное влияние лишь на популяцию в целом, повышая жизнеспособность вида). К трансмиссивным относят плазмиды, инициирующие свойства доноров у клеток-хозяев. При этом последние получают новое качество - возможность конъюгировать с клетками-реципиентами и отдавать им свои плазмиды. Клетки-реципиенты, приобретая во время конъюгации плазмиды, сами превращаются в доноров.[ ...]
Отсутствие адсорбции не исчерпывает разнообразия вариантов взаимодействия бактериальных вирусов и микробных клеток. Они иллюстрируют лишь одну сторону этого явления, а именно проявление клеточных защитных механизмов, фенотипически (по критерию отсутствия роста) иммитирующих рестрикцию. Однако, существует и другой вариант взаимодействия клетка-бактериофаг, который может иммитировать отсутствие СХС. Примерами таких механизмов является синтез ингибиторов и метилаз кодируемых фаговыми генами, защищающих вирусную ДНК от действия рестриктаз II типа.[ ...]
Механизм обеззараживающего действия хлора связан с нарушением обмена веществ бактериальной клетки в процессе дезинфекции воды. При этом выявлено влияние на ферментную активность бактерий, в частности, на дегидрогеназы, катализирующие окислительно-восстановительные реакции в бактериальной клетке. А. М. Ски-дальской (1969) было изучено влияние хлора на процесс декарбоксилирования аминокислот бактерий, протекающий в присутствии строго специфичных ферментов-декарбоксилаз, а также был определен нуклеотидный состав ДНК кишечной палочки после окончания процесса обеззараживания при различных уровнях бактерицидного эффекта.[ ...]
Бактериофаги Т-группы имеют форму барабанных палочек размером 100 х 25 нм. Их геном представлен ДНК. Они являются вирулентными фагами, т. к. после инфицирования ими бактериальных клеток последние лизируются с освобождением большого количества вновь синтезированных фаговых частиц.[ ...]
Плазмиды бактерий - это генетические структуры, находящиеся в цитоплазме и представляющие собой молекулы ДНК размером от 2250 до 400 ООО пар азотистых оснований. Они существуют обособленно от хромосом в количестве от одной до нескольких десятков копий на одну бактериальную клетку.[ ...]
Штамм Pseu.dom.onas вуг1 ае ри. рка8еоИсо1а обладает плазмидой длиной 150 тыс. п. н., которая может реплицироваться автономно, а может интегрироваться в бактериальную хромосому. Последующее неточное вырезание позволило получить семейство плазмид длиной от 35 до 270 тыс. п. н., некоторые из них содержали большие сегменты хромосомной ДНК .[ ...]
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикции-модификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внухриклеточно, т. е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой причине чужеродная ДНК из-за отсутствия в ней метальных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нуклеотидов. Можно сказать, что таким образом бактерии защищают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).[ ...]
Наследственные свойства бактерий или отдельные признаки закодированы в единицах наследственности - генах, линейно расположенных в хромосоме вдоль нити ДНК. Следовательно, ген является фрагментом нити ДНК- Каждому признаку соответствует определенный ген, а часто еще меньший отрезок ДНК - кодон. Иначе говоря, в нити ДНК в линейном порядке расположена информация обо всех свойствах бактерий. При этом у бактерий есть еще одна особенность. В ядрах эукариотов содержится обычно несколько хромосом, число их в ядре постоянно у каждого вида. Нуклеоид бактерий содержит лишь одно кольцо из нити ДНК, т. е. одну хромосому. Однако запасом информации, заключенным в одной хромосоме или в кольцеобразно сомкнувшейся двунитчатой спирали ДНК, сумма наследственных признаков бактериальной клетки не исчерпывается. Плазмиды содержат ДНК, также несущую генетическую информацию, передаваемую от материнской клетки к дочерней.[ ...]
Мутации - это изменения в генном аппарате клетки, которые сопровождаются изменениями контролируемых этими генами признаков. Различают макро- и микроповреждения ДНК, ведущие к изменению свойств клетки. Макроизменения, а именно: выпадение участка ДНК (деления), перемещение отдельного участка (транслокация) или поворот определенного участка молекулы на 180° (инверсия) -у бактерий наблюдаются сравнительно редко Гораздо более характерны для них микроповреждения, или точечные мутации, т. е. качественные изменения в отдельных генах, например замена пары азотистых оснований. Мутации бывают прямые и обратные, или реверсивные. Прямые - это мутации организмов дикого типа, например утрата способности самостоятельно синтезировать факторы роста, т. е. переход от прото- к ауксотрофности. Обратные мутации представляют собой возвращение, или реверсию, к дикому типу. Способность к реверсии характерна для точечных мутаций. В результате мутаций изменяются такие важнейшие признаки, как способность самостоятельно синтезировать аминокислоты и витамины (ауксотрофные мутанты), способность к образованию ферментов. Эти мутации называют биохимическими. Хорошо известны также мутации, ведущие к изменению чувствительности к антибиотикам и другим антимикробным веществам. По происхождению мутации разделяют на спонтанные и индуцированные. Спонтанные возникают самопроизвольно без вмешательства человека и носят случайный характер. Частота таких мутаций очень низка и составляет от 1 X Ю“4 ло 1 X 10-10. Индуцированные возникают при воздействии на микроорганизмы физических или химических мутагенных факторов. К физическим факторам, обладающим мутагенным действием, относятся ультрафиолетовое и ионизирующие излучения, а также температура. Химическими мутагенами являются ряд соединений и среди них наиболее активны так называемые супермутагены. В природных условиях и эксперименте изменения в составе бактериальных популяций могут возникать в результате действия двух факторов - мутаций и автоселекции, происходящей в результате адаптации некоторых мутантов к условиям среды обитания. Такой процесс, очевидно, наблюдается в среде, где преобладающим источником питания является синтетическое вещество, например, ПАВ или капролактам.[ ...]
Одиночная клетка E. coli окружена трехслойной клеточной оболочкой толщиной порядка 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде, примерно, 2 х Ю 1Я г белка, 6 х 10 16 г ДНК и 2 х 10 14 г РНК (в основном ри-босомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10“® М, тогда как других - порядка 2 х 10"4 М (от 10 до 200 000 молекул на клетку).[ ...]
У одноклеточных организмов половое размножение существует в нескольких формах. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.[ ...]
Бактерии: прокариоты («доядерные») одноклеточные организмы. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированные для фотосинтеза) и митохондрии (специализированные для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.[ ...]
Крайне малые размеры клеток являются характерной, но не главной особенностью бактерий. Все бактерии представлены особым типом клеток, лишенных истинного ядра, окруженного ядерной мембраной. Аналогом ядра у бактерий является нуклеоид - ДНК-содержащая плазма, не отграниченная от цитоплазмы мембраной. Кроме того, для бактериальных клеток характерны отсутствие митохондрий, хлоропла-стов, а также особое строение и состав мембранных структур и клеточных стенок. Организмы, в клетках которых отсутствует истинное ядро, называются прокариотами (доядер-ными) или протоцитами (т. е. организмами с примитивной организацией клеток).[ ...]
Микоплазменные клетки имеют овальную форму, а их размеры составляют около 0,1-0,25 нм в диаметре (рис. 43). Для них характерно наличие тонкой наружной плазматической мембраны (толщина - около 8 нм), которая окружает цитоплазму, содержащую молекулу ДНК, достаточную для кодирования около 800 разных белков, РНК разных типов, рибосом диаметром порядка 20 нм. В их цитоплазме содержатся различные включения в виде белков, гранул липидов и других соединений. Из-за недостаточной жесткости клеточкой мембраны микоплазмы проходят через бактериальные фильтры.[ ...]
Установлено, что на рибосомах происходят связывание активированных аминокислот и укладка их в полипептидную цепь в соответствии с генетической информацией, полученной из ядра через информационную (матричную) РНК (мРНК), которая как бы считывает соответствующую информацию с ДНК и передает ее на рибосомы. Целый ряд белков синтезирован на изолированных рибосомах и при этом отмечено включение в них меченых аминокислот. Роль матрицы в белковом синтезе выполняет мРНК, которая прикрепляется к рибосоме. На поверхности последней происходит взаимодействие между комплексом аминокислот, транспортной РНК, несущей очередную аминокислоту, и нуклеотидной последовательностью информационной РНК, которая функционирует на рибосоме однократно и после синтеза полипептидной цепи распадается, а вновь синтезированный белок накапливается в рибосомах. В бактериальной клетке при периоде регенерации 90 мин скорость кругооборота мРНК достигает 4-6 с.[ ...]
Цитоплазма представляет собой коллоидный раствор, дисперсной фазой которого являются сложные белковые соединения и вещества, близкие к жирам, а дисперсионной средой - вода. У некоторых форм бактерий в цитоплазме содержатся включения - капельки жира, серы, гликогена и др. Постоянными составляющими бактериальных клеток являются особые выросты цитоплазматической мембраны - мезосомы, в которых содержатся ферментные окислительно-восстановительные системы. В этих образованиях идут в основном процессы, связанные с дыханием бактерий. В мелких включениях - рибосомах, содержащих рибонуклеиновую кислоту, осуществляется биосинтез белка. Большинство видов бактерий не имеет обособленного ядра. Ядерное вещество, представленное ДНК, у них не отделено от цитоплазмы и образует нуклеоид. Транспортировка веществ, необходимых для жизнедеятельности клетки, и отвод продуктов обмена осуществляется по особым каналам и полостям, отделенным от цитоплазмы мембраной, имеющей такое же строение, как и цитоплазматическая. Это структурное образование называется эндоплазматической сетью (ретикулум).[ ...]
Представление об изменчивости и наследственности бактерий нельзя составить без знания некоторых положений молекулярной генетики прокариотической клетки. В основе процессов приспособления микробных культур к изменяющимся экологическим условиям лежат изменчивость и наследственность, являющиеся разделами генетики бактерий. При изложении цитологии бактериальной клетки уже рассматривалась структура ДНК и РНК и их роль в жизни клетки. Характерное строение ДНК сохраняется у каждого вида и передается потомству из поколения в поколение, как и другие признаки. ДНК бактерий представляет собой двунитчатую спираль, замыкающуюся в кольцо. Кольчатая нить ДНК бактерий, расположенная в ну-клеоиде, не содержит белка. Такое кольцо ДНК соответствует хромосоме эукариотической клетки. Известно, что в хромосоме эукариотических клеток, кроме ДНК, всегда содержится белковый компонент. Отсюда следует, что понятие хромосомы у эукариотов несколько отлично от понятия хромосомы бактерий. Нить ДНК, представляющая собой хромосому бактерий, разумеется, у разных видов различается. Сахарофосфатный компонент ДНК У всех видов бактерий одинаков; расположение азотистых оснований и их комбинация, напротив, различаются у разных видов.[ ...]
Все возрастающее беспорядочное применение антибиотиков в животноводстве, которые используются в малых дозах как стимуляторы роста, а также в качестве превентивной меры против вызванных стрессом желудочно-кишечных расстройств у животных на фермах, приводит к все более широкому распространению в микробных популяциях R-фактора устойчивости к антибиотикам, передающегося от одной бактериальной клетки к другой при конъюгации. Передача происходит через плазмиду, которая представляет собой кольцевую экстрахромосомную ДНК, способную к репликации.[ ...]
В противоположность вирулентным фагам, известны так называемые фаги умеренного действия, или просто умеренные фаги. Типичным представителем таких фагов является фаг X, который тоже использовался и используется в качестве экспериментальной модели для выяснения многих вопросов молекулярной генетики. Фагу X присущи два важных свойства. Подобно вирулентным фагам он может инфицировать бактериальные клетки, размножаться вегетативно, продуцируя в клетках сотни копий и лизировать клетки С освобождением зрелых фагочастиц. Однако ДНК этого фага может включаться в бактериальную хромосому, превращаясь в профаг. При этом происходит так называемая лизогенизация бактерий, а бактерии, содержащие профаг, называют лизогенными. Лизогенные бактериальные клетки могут обладать профагом бесконечно долгое время, при этом не лизируясь. Лизис с освобождением новых фагочастиц отмечается после воздействия на лизогенные бактерии какого-либо фактора, например УФ-излучения, которое индуцирует развитие профага в фаг. Изучение лизогенных бактерий позволило получить ряд новых данных о роли разных белков в действии фаговых генов.[ ...]
Геном хлоропластов ряда высших растений состоит из 120 генов. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и по функциям. В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены. Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.[ ...]
Современные схемы, иллюстрирующие работу генов, построены на основании логического анализа экспериментальных данных, полученных с помощью биохимических и генетических методов. Применение тонких электронно-микроскопических методов позволяет в буквальном смысле слова увидеть работу наследственного аппарата клетки. В последнее время получены электронно-микроскопические снимки, на которых видно, как на матрице бактериальной ДНК, в тех участках, где к ДНК прикреплены молекулы РНК-полимеразы (фермента, катализирующего транскрипцию ДНК в РНК), происходит синтез молекул и-РНК. Нити и-РНК, расположенные перпендикулярно к линейной молекуле ДНК, продвигаются вдоль матрицы и увеличиваются в длине. По мере удлинения нитей РНК к ним присоединяются рибосомы, которые, продвигаясь, в свою очередь, вдоль нити РНК по направлению к ДНК, ведут синтез белка.[ ...]
Трансдукция - это перенос генетического материала от бактерии-донора к бактерии-реципиенту с помощью фага. Впервые явление трансдукции было открыто в 1951 г. Ледербергом с сотрудниками у Salmonella typhimurium. Сейчас различают неспецифическую и специфическую трансдукции. При неспецифической трансдукции возможен перенос фагом любого признака от бактерии-донора к бактерии-реципиенту. Перенос осуществляется только умеренными (невирулентными) фагами. Умеренные фаги способны заражать бактерии, однако не размножаются в них и не вызывают лизиса, а включаются в ДНК бактериальной клетки и в таком неинфекционном состоянии в виде так называемого профага передаются от клетки к клетке при размножении. Культуры бактерий, содержащие профаг, называются лизогенными. В этих культурах с небольшой частотой (в одной из 102 - 105 клеток) наблюдается спонтанное размножение фага и происходит лизис клетки с освобождением фаговых частиц, обнаруживаемых с помощью бактерий-индикаторов, для которых такой фаг вирулентен.[ ...]
Опыты проводили на трехкамерной ячейке, состоящей из центральной рабочей и двух электродных камер. В рабочую камеру размером 25 X 7 X 37 мм (длина X ширина X высота), отделенную от электродных целлофановыми мембранами, помещали 750 мг ваты. Через нее снизу вверх подавали с постоянной скоростью исходный раствор исследуемых веществ. За содержанием соединений в исходных, подаваемых в рабочую камеру растворах (С0), и в растворах, выходящих из камеры (Ci), следили по максимумам поглощения белков и нуклеиновых кислот в диапазоне волновых чисел (35,5-38) X Ю3 см-1 с помощью УФ-спектрофотометра Specord UV-VIS. Электродные камеры заполняли гранулированным активированным углем и через них отдельным протоком пропускали дистиллированную воду.
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.

Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.
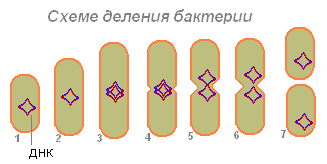
После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Бактерии – прокариотические микроорганизмы, генетический материал которых в основном представлен единственной кольцевой двухцепочечной ДНК, называемой генетиками хромосомой. В относительно редких случаях хромосома представлена линейной молекулой ДНК.
Размер этой ДНК намного превышает размер самой бактериальной клетки. Так, например, у E. coli протяженность хромосомной ДНК равна 1300 мкм (1,3 мм – 4,6 х 10 6 п.н.), а размер клетки 1,1-1,5 х 2,0-6,0 мкм. Причем ДНК не заполняет всю клетку, а содержится только в ограниченной области, составляющей, весьма приблизительно, одну треть объема клетки.
   |
Рис.1. Бактериальный геном и схема уровней его компактизации.
Отсюда следует, что ДНК существует в клетке в высокоупорядоченном (сконденсированном) состоянии в виде компактной структуры. Эта структура, отдаленно напоминая ядра эукариот, получила название нуклеоид и видна в микроскопе только после специфичных для ДНК окрасок (рис.1). В электронном микроскопе она выглядит как образование, состоящее из многочисленных петель, отходящих от плотной центральной области. Образование большого числа (до 140 на геном) петель, называемых доменами , является одним из уровней компактизации ДНК. Каждый домен закреплен у основания молекулой РНК и состоит примерно из 40 т.п.н. ДНК петель находится не в виде свободно вытянутого дуплекса, а имеет второй уровень компактизации за счет скручивания в сверхспиральные образования с помощью связи с белками HLP.
Эти белки имеют небольшой размер, обладают сильноосновными свойствами и прочно связываются с ДНК. По аминокислотному составу они напоминают гистоны эукариот.
Нуклеоид не отделен от цитоплазмы ядерной мембраной и прикреплен к мезосомам – специфическим впячиваниям цитоплазматической мембраны внутрь клетки. Связь ДНК со специфическим участком мембраны необходима для функционирования генома.
Кольцевая молекула ДНК бактерий (хромосома) представляет самореплицирующуюся генетическую молекулу – репликон . Репликация начинается с точки инициации репликации (ori – orign) , локализующейся, как правило, в участке прикрепления ДНК к мембране. От точки инициации репликация происходит последовательно, двунаправленно, по полуконсервативному механизму. Заканчивается репликация в районе терминации репликации (ter) , расположенном на участке кольцевой ДНК, противоположном точке начала репликации (рис.2).
Рис. 3. Распределение дочерних копий ДНК и деление клетки бактерий.
Число хромосом в одной клетке бактерий зависит от стадии развития и физиологических условий роста культуры. В логарифмической стадии роста у E. coli на 1 нуклеоид приходится 2,8 ДНК эквивалентов одного генома, вследствие замедленной сегрегации двух дочерних хромосом, или реинициации новых циклов репликации ДНК еще до деления клетки (рис.4).
 |
Рис.4. Число хромосом в клетке в стационарной (А) и логарифмической (Б) стадиях роста культуры.
У некоторых бактерий клетки в норме содержат не одну, а много хромосом. Они могут формировать один или несколько нуклеоидов. Также наблюдается зависимость содержания ДНК в клетке от ее размеров, хотя это не означает соответствующего изменения объема генетической информации.
Для бактериальной ДНК характерна высокая плотность генов (1 ген на 1тпн). ДНК, кодирующая белки, составляет около 85-90% всей ДНК. Средний размер ДНК-последовательностей между генами – только 110-125 п.н. Некодирующая бактериальная ДНК занимает менее 1%, и она обычно представлена в виде транспозонов. Так, в ДНК штамма Escherichia coli K12 линии MG 1655 найдена 41 копия различных транспозонов (IS), которые участвуют в процессах внедрения и исключения плазмид. Многие фаги, исключаясь из генома бактерии не полностью, оставляют там в качестве следа некоторые свои гены. Эти остатки, не способные к самостоятельному перемещению и развитию, называют "криптическими" фагами.
Интроны встречаются в бактериальных геномах крайне редко. Имеются случаи перекрывания генов, где один ген находится внутри другого на той же нити ДНК. Для бактериальных геномов характерны опероны: у Е. coli 27% предсказанных транскрипционных единиц являются оперонами.
В клетке бактерий могут содержаться и другие репликоны, способные существовать отдельно от бактериальной хромосомы. Их называют плазмидами . Плазмиды представляют собой кольцевые (у некоторых видов линейные) молекулы двухцепочечной ДНК различных размеров от 1000 п.н. до почти трети величины самой бактериальной хромосомы. Количество и спектр плазмид в клетках бактерий может варьировать. Часто наблюдаются различия по спектру плазмид даже между клетками разных штаммов одного и того же вида бактерий. Некоторые плазмиды могут встраиваться в бактериальную хромосому, составляя при этом часть репликона бактерии, и могут вновь исключаться из нее, восстанавливая форму автономного репликона. Такие плазмиды называют эписомами .
В генетический материал бактерий могут быть включены и профаги.
Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
ДНК (дезоксирибонуклеиновая кислота) - своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение. ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни как человека, так и любого др. организхма. Искусственное или естественное воздействие внешней среды способны лишь в незначительной степени повлиять на общую выраженность отдельных генетических признаков или сказаться на развитии запрограммированных процессов.
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С ) и фосфатной (Ф ) группы (фосфодиэфирные связи).

Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т ), гуанин — только с цитозином (Г-Ц ). Именно эти пары и составляют «перекладины» винтовой "лестницы" ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
- Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной.
- Двойная спираль «расстегивается» на две ветви, когда ферменты разрушают связь между базовыми парами химических соединений.
- Каждая ветвь является элементом новой ДНК. Новые базовые пары соединяются в той же последовательности, что и в родительской ветви.
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Рис. 4 . Азотистые основания: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты - это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
НУКЛЕОТИДЫ состоят из азотистого основания , соединенного с пятиуглеродным углеводом (пентозой) - дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H 2 PO 3 -).
Азотистые основания бывают двух типов: пиримидиновые основания - урацил (только в РНК), цитозин и тимин, пуриновые основания - аденин и гуанин.

Рис. 5. Структура нуклеотидов (слева), расположение нуклеотида в ДНК (снизу) и типы азотистых оснований (справа): пиримидиновые и пуриновые

Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль . Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей . Аденин всегда соединяется с тимином, а цитозин - с гуанином. Это называется правилом комплементарности .
Правило комплементарности:
| A-T G-C |
Например, если нам дана цепь ДНК, имеющая последовательность
3’- ATGTCCTAGCTGCTCG - 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении - от 5’-конца к 3’-концу:
5’- TACAGGATCGACGAGC- 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ ДНК
Репликация ДНК - это процесс удвоения молекулы ДНК путем матричного синтеза. В большинстве случаев естественной репликации ДНК праймером для синтеза ДНК является короткий фрагмент (создаваемый заново). Такой рибонуклеотидный праймер создается ферментом праймазой (ДНК-праймаза у прокариот, ДНК-полимераза у эукариот), и впоследствии заменяется дезоксирибонуклеотидами полимеразой, выполняющей в норме функции репарации (исправления химических повреждений и разрывов в молекле ДНК).
Репликация происходит по полуконсервативному механизму. Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез - это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:
1) ДНК-топоизомераза, располагаясь перед вилкой репликации, разрезает ДНК для того, чтобы облегчить ее расплетание и раскручивание.
2) ДНК-хеликаза вслед за топоизомеразой влияет на процесс «расплетения» спирали ДНК.
3) ДНК-связывающие белки осуществляют связывание нитей ДНК, а также проводят их стабилизацию, не допуская их прилипания друг к другу.
4) ДНК-полимераза δ
(дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез
ведущей
цепи
дочерней ДНК в направлении 5"→3" на матрице
материнскойнити ДНК по направлению от ее 3"-конца к 5"-концу (скорость до 100 пар нуклеотидов в секунду). Этим события на данной материнской
нити ДНК ограничиваются.

Рис. 9. Схематическое изображение процесса репликации ДНК: (1) Отстающая цепь (запаздывающая нить), (2) Ведущая цепь (лидирующая нить), (3) ДНК-полимераза α (Polα ), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза δ (Polδ ), (9) Хеликаза, (10) Однонитевые ДНК-связывающие белки, (11) Топоизомераза.
Далее описан синтез отстающей цепи дочерней ДНК (см. Схему репликативной вилки и функции ферментов репликации)
Нагляднее о репликации ДНК см.
5) Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (альфа) и в направлении 5"→3" синтезирует праймер (РНК-затравку) - последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
Вместо
ДНК-полимеразы
α
к 3"-концу праймера присоединяется
ДНК-полимераза
ε
.
6)
ДНК-полимераза
ε
(эпсилон)
как бы продолжает удлинять праймер, но в качестве субстрата встраивает
дезоксирибонуклеотиды
(в количестве 150-200 нуклеотидов). В результате образуется цельная нить из двух частей -
РНК
(т.е. праймер) и
ДНК
.
ДНК-полимераза
ε
работает до тех пор, пока не встретит праймер предыдущего
фрагмента Оказаки
(синтезированный чуть ранее). После этого данный фермент удаляется с цепи.
7) ДНК-полимераза β (бета) встает вместо ДНК-полимеразы ε , движется в том же направлении (5"→3") и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид (еще более ранее синтезированный ДНК-полимеразой ε ). Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи.
В результате на матрице материнской нити "лежит" фрагмент дочерней ДНК. Он называется
фрагмент Оказаки
.
8) ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки , т.е. 5"-конца отрезка, синтезированного ДНК-полимеразой ε , и 3"-конца цепи, встроенного ДНК-полимеразой β .
СТРОЕНИЕ РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК, РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом . Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Однако в отличие от ДНК, РНК обычно имеет не две цепи, а одну. Пентоза в РНК представлена рибозой, а не дезоксирибозой (у рибозы присутствует дополнительная гидроксильная группа на втором атоме углевода). Наконец, ДНК отличается от РНК по составу азотистых оснований: вместо тимина (Т ) в РНК представлен урацил (U ) , который также комплементарен аденину.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией , то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами - РНК-полимеразами .
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК - эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ - 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.
![]()
Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’- ATGTCCTAGCTGCTCG - 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’- TACAGGATCGACGAGC- 3’,
а синтезируемая с нее РНК - последовательность
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка
на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:

Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
ГЕНЕТИЧЕСКИЙ КОД
Генетический код - способ кодирования аминокислотной последовательности белков с помощью последовательности нуклеотидов. Каждая аминокислота кодируется последовательностью из трех нуклеотидов - кодоном или триплетом.
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5" к 3" концу мРНК.
Таблица 1. Стандартный генетический код
|
1-е
ние |
2-е основание |
3-е
ние |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Tyr/Y) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
U C A |
U A A |
Стоп-кодон** |
U G A |
Стоп-кодон** |
A |
||
|
U U G |
U C G |
U A G |
Стоп-кодон** |
U G G |
(Trp/W) |
G |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(His/H) |
C G U |
(Arg/R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
A |
||||
|
A U G |
(Met/M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu/E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
- *Триплет AUG , также кодирующий метионин, называется старт-кодоном . С этого кодона начинается синтез молекулы белка. Таким образом, во время синтеза белка, первой аминокислотой в последовательности всегда будет метионин.
- **Триплеты UAA , UAG и UGA называются стоп-кодонами и не кодируют ни одной аминокислоты. На этих последовательностях синтез белка прекращается.
Свойства генетического кода
1. Триплетность . Каждая аминокислота кодируется последовательностью из трех нуклеотидов - триплетом или кодоном.
2. Непрерывность . Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.
3. Неперекрываемость . Один нуклеотид не может входить одновременно в два триплета.
4. Однозначность . Один кодон может кодировать только одну аминокислоту.

5. Вырожденность . Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность . Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’- CCGATTGCACGTCGATCGTATA - 5’.
Матричная цепь будет иметь последовательность:
5’- GGCTAACGTGCAGCTAGCATAT - 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’- CCGAUUGCACGUCGAUCGUAUA - 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’- AUAUGCUAGCUGCACGUUAGCC - 3’.
Теперь найдем старт-кодон AUG:
5’- AUAUG CUAGCUGCACGUUAGCC - 3’.
Разделим последовательность на триплеты:
звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК - на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
(общие понятия)
Геном - совокупность всех генов организма; его полный хромосомный набор.
Термин "геном" был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими ("избыточными") последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.
В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации ), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент» . Позднее эта концепция была расширена до определения «один ген — один полипептид» , поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
На рис. 14 показана схема того, как триплеты нуклеотидов в ДНК определяют полипептид - аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками(так называемыми интронами ).
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена , кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов ). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
 Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру - нуклеоид. Хромосома прокариота Escherichia coli , чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Бактерия E. coli
имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli
приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами
(рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
См. также: Равин Н.В., Шестаков С.В. Геном прокариот // Вавиловский журнал генетики и селекции, 2013. Т. 17. № 4/2. С. 972-984.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов
|
Общая ДНК, п.н. |
Число хромосом* |
Примерное число генов |
|
|
Escherichia coli (бактерия) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans (нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana (растение) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
20 000 |
|
|
Oryza sativa (рис) |
480 000 000 |
57 000 |
|
|
Mus musculus (мышь) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens (человек) |
3 070 128 600 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам
* Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом. Диплоидный
набор хромосом (от греч. diploos- двойной и eidos- вид) - двойной набор хромосом (2n), каждая из которых имеет себе гомологичную.
**Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
***Для самок с двумя Х хромосомами. У самцов есть Х хромосома, но нет Y, т. е. всего 11 хромосом.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке E. coli (табл. 2). Клетки плодовой мушки Drosophila , классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки E. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2n ) зависит от вида организма (табл. 2).
Например, в соматической клетке человека 46 хромосом (рис. 17 ). Каждая хромосома эукариотической клетки, как показано на рис. 17, а , содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.

Рис. 17. Хромосомы эукариот. а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.
Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или Х и Х), получится последовательность длиной около одного метра. Прим.: У всех млекопитающих и других организмов с гетерогаметным мужским полом, у самок две X-хромосомы (XX), а у самцов — одна X-хромосома и одна Y-хромосома (XY).
Большинство клеток человека , поэтому общая длина ДНК таких клеток около 2м. У взрослого человека примерно 10 14 клеток, таким образом, общая длина всех молекул ДНК составляет 2・10 11 км. Для сравнения, окружность Земли — 4・10 4 км, а расстояние от Земли до Солнца — 1,5・10 8 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК, — это митохондрии и хлоропласты. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл. Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК.
СТРОЕНИЕ ГЕНОВ
Рассмотрим строение гена у прокариот и эукариот, их сходства и различия. Несмотря на то, что ген — это участок ДНК, кодирующий всего один белок или РНК, кроме непосредственно кодирующей части, он также включает в себя регуляторные и иные структурные элементы, имеющие разное строение у прокариот и эукариот.
Кодирующая последовательность - основная структурно-функциональная единица гена, именно в ней находятся триплеты нуклеотидов, кодирующие аминокислотную последовательность. Она начинается со старт-кодона и заканчивается стоп-кодоном.
До и после кодирующей последовательности находятся нетранслируемые 5’- и 3’-последовательности . Они выполняют регуляторные и вспомогательные функции, например, обеспечивают посадку рибосомы на и-РНК.
Нетранслируемые и кодирующая последовательности составлют единицу транскрипции - транскрибируемый участок ДНК, то есть участок ДНК, с которого происходит синтез и-РНК.
Терминатор - нетранскрибируемый участок ДНК в конце гена, на котором останавливается синтез РНК.
В начале гена находится регуляторная область , включающая в себя промотор и оператор .
Промотор - последовательность, с которой связывается полимераза в процессе инициации транскрипции. Оператор - это область, с которой могут связываться специальные белки - репрессоры , которые могут уменьшать активность синтеза РНК с этого гена - иначе говоря, уменьшать его экспрессию .
Строение генов у прокариот
Общий план строения генов у прокариот и эукариот не отличается - и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор. Однако организация генов у прокариот и эукариот отличается.

Рис. 18. Схема строения гена у прокариот (бактерий) - изображение увеличивается
В начале и в конце оперона есть единые регуляторные области для нескольких структурных генов. С транскрибируемого участка оперона считывается одна молекула и-РНК, которая содержит несколько кодирующих последовательностей, в каждой из которых есть свой старт- и стоп-кодон. С каждого из таких участков с интезируется один белок. Таким образом, с одной молекулы и-РНК синтезируется несколько молекул белка.
Для прокариот характерно объединение нескольких генов в единую функциональную единицу - оперон . Работу оперона могут регулировать другие гены, которые могут быть заметно удалены от самого оперона - регуляторы . Белок, транслируемый с этого гена называется репрессор . Он связывается с оператором оперона, регулируя экспрессию сразу всех генов, в нем содержащихся.
Для прокариот также характерно явление сопряжения транскрипции и трансляции .
![]()
Рис. 19 Явление сопряжения транскрипции и трансляции у прокариот - изображение увеличивается
Такое сопряжение не встречается у эукариот из-за наличия у них ядерной оболочки, отделяющей цитоплазму, где происходит трансляция, от генетического материала, на котором происходит транскрипция. У прокариот во время синтеза РНК на матрице ДНК с синтезируемой молекулой РНК может сразу связываться рибосома. Таким образом, трансляция начинается еще до завершения транскрипции. Более того, с одной молекулой РНК может одновременно связываться несколько рибосом, синтезируя сразу несколько молекул одного белка.
Строение генов у эукариот
Гены и хромосомы эукариот очень сложно организованы
У бактерий многих видов всего одна хромосома, и почти во всех случаях в каждой хромосоме присутствует по одной копии каждого гена. Лишь немногие гены, например гены рРНК, содержатся в нескольких копиях. Гены и регуляторные последовательности составляют практически весь геном прокариот. Более того, почти каждый ген строго соответствует аминокислотной последовательности (или последовательности РНК), которую он кодирует (рис. 14).
Структурная и функциональная организация генов эукариот гораздо сложнее. Исследование хромосом эукариот, а позднее секвенирование полных последовательностей геномов эукариот принесло много сюрпризов. Многие, если не большинство, генов эукариот обладают интересной особенностью: их нуклеотидные последовательности содержат один или несколько участков ДНК, в которых не кодируется аминокислотная последовательность полипептидного продукта. Такие нетранслируемые вставки нарушают прямое соответствие между нуклеотидной последовательностью гена и аминокислотной последовательностью кодируемого полипептида. Эти нетранслируемые сегменты в составе генов называют интронами , или встроенными последовательностями , а кодирующие сегменты — экзонами . У прокариот лишь немногие гены содержат интроны.
Итак, у эукариот практически не встречается объединение генов в опероны, и кодирующая последовательность гена эукариот чаще всего разделена на транслируемые участки - экзоны , и нетранслируемые участки - интроны.
В большинстве случаев функция интронов не установлена. В целом, лишь около 1,5% ДНК человека являются ≪кодирующими≫, т. е. несут информацию о белках или РНК. Однако с учетом крупных интронов получается, что ДНК человека на 30% состоит из генов. Поскольку гены составляют относительно небольшую долю в геноме человека, значительная часть ДНК остается неучтенной.

Рис. 16. Схема строение гена у эукариот - изображение увеличивается
С каждого гена сначала синтезируется незрелая, или пре-РНК, которая содержит в себе как интроны, так и экзоны.
После этого проходит процесс сплайсинга, в результате которого интронные участки вырезаются, и образуется зрелая иРНК, с которой может быть синтезирован белок.

Рис. 20. Процесс альтернативного сплайсинга - изображение увеличивается
Такая организация генов позволяет, например, осуществить , когда с одного гена могут быть синтезированы разные формы белка, за счет того, что в процессе сплайсинга экзоны могут сшиваться в разных последовательностях.

Рис. 21. Отличия в строении генов прокариот и эукариот - изображение увеличивается
МУТАЦИИ И МУТАГЕНЕЗ
Мутацией называется стойкое изменение генотипа, то есть изменение нуклеотидной последовательности.
Процесс, который приводит к возникновению мутаций называется мутагенезом , а организм, все клетки которого несут одну и ту же мутацию — мутантом .
Мутационная теория была впервые сформулирована Гуго де Фризом в 1903 году. Современный ее вариант включает в себя следующие положения:
1. Мутации возникают внезапно, скачкообразно.
2. Мутации передаются из поколения в поколение.
3. Мутации могут быть полезными, вредными или нейтральными, доминантными или рецессивными.
4. Вероятность обнаружения мутаций зависит от числа исследованных особей.
5. Сходные мутации могут возникать повторно.
6. Мутации не направленны.
Мутации могут возникать под действием различных факторов. Различают мутации, возникшие под действием мутагенных воздействий : физических (например, ультрафиолета или радиации), химических (например, колхицина или активных форм кислорода) и биологических (например, вирусов). Также мутации могут быть вызваны ошибками репликации .
В зависимости от условий появления мутации подразделяют на спонтанные — то есть мутации, возникшие в нормальных условиях, и индуцированые — то есть мутации, которые возникли при особых условиях.
Мутации могут возникать не только в ядерной ДНК, но и, например, в ДНК митохондрий или пластид. Соответственно, мы можем выделять ядерные и цитоплазматические мутации.
В результате возникновения мутаций часто могут появляться новые аллели. Если мутантный аллель подавляет действие нормального, мутация называется доминантной . Если нормальный аллель подавляет мутантный, такая мутация называется рецессивной . Большинство мутаций, приводящих к возникновению новых аллелей являются рецессивными.
По эффекту выделяют мутации адаптивные , приводящие к повышению приспособленности организма к среде, нейтральные , не влияющие на выживаемость, вредные , понижающие приспособленность организмов к условиям среды и летальные , приводящие к смерти организма на ранних стадиях развития.
По последствиям выделяются мутации, приводящие к потери функции белка , мутации, приводящие к возникновению у белка новой функции , а также мутации, которые изменяют дозу гена , и, соответственно, дозу белка синтезируемого с него.
Мутация может возникнуть к любой клетке организма. Если мутация возникает в половой клетке, она называется герминативной (герминальной, или генеративной). Такие мутации не проявляются у того организма, у которого они появились, но приводят к появлению мутантов в потомстве и передаются по наследству, поэтому они важны для генетики и эволюции. Если мутация возникает в любой другой клетке, она называется соматической . Такая мутация может в той или иной степени проявляться у того организма, у которого она возникла, например, приводить к образованию раковых опухолей. Однако такая мутация не передается по наследству и не влияет на потомков.
Мутации могут затрагивать разные по размеру участки генома. Выделяют генные , хромосомные и геномные мутации.
Генные мутации
Мутации, которые возникают в масштабе меньшем, чем один ген, называются генными , или точечными (точковыми) . Такие мутации приводят к изменению одного и нескольких нуклеотидов в последовательности. Среди генных мутаций выделяют замены , приводящие к замене одного нуклеотида на другой, делеции , приводящие к выпадению одного из нуклеотидов, инсерции , приводящие к добавлению лишнего нуклеотида в последовательность.

Рис. 23. Генные (точечные) мутации
По механизму воздействия на белок, генные мутации делят на: синонимичные , которые (в результате вырожденности генетического кода) не приводят к изменению аминокислотного состава белкового продукта, миссенс-мутации , которые приводят к замене одной аминокислоты на другую и могут влиять на структуру синтезируемого белка, хотя часто они оказываются незначительными, нонсенс-мутации , приводящие к замене кодирующего кодона на стоп-кодон, мутации, приводящие к нарушению сплайсинга:

Рис. 24. Схемы мутаций
Также по механизму воздействия на белок выделяют мутации, приводящие к сдвигу рамки
считывания
, например, инсерции и делеции. Такие мутации, как и нонсенс-мутации, хоть и возникают в одной точке гена, часто воздействуют на всю структуру белка, что может привести к полному изменению его структуры.

Рис. 29. Хромосома до и после дупликации
Геномные мутации
Наконец, геномные мутации затрагивают весь геном целиком, то есть меняется количество хромосом. Выделяют полиплоидии — увеличение плоидности клетки, и анеуплоидии, то есть изменение количества хромосом, например, трисомии (наличие у одной из хромосом дополнительного гомолога) и моносомии (отсутствие у хромосомы гомолога).
Видео по теме ДНК
РЕПЛИКАЦИЯ ДНК, КОДИРОВАНИЕ РНК, СИНТЕЗ БЕЛКА


